
Yesterday saw the publication of my most recent paper. This one is a first for me in several ways. It is my first description of a new species based on specimens from the museum collection which I now oversee, and also, the first pinniped I get to properly describe.
The new paper came out in the open access journal PeerJ, that means you can download the whole article for free here!! In it I describe a new species of basal otariid (this is the group of pinnipeds that include fur seals and sea lions). The specimen comes from a very productive locality near the Upper Oso Reservoir in Orange County, California (see map below). Staff and colleagues from the Natural History Museum of Los Angeles (NHMLA) collected at this site in the 1970s (you can see part of the site in this video, starting around minute 4:46). However, not much has been published so far.
 |
| FIGURE 1. Location of the study site (Upper Oso Dam) in Southern California. |
At the locality, which is called Upper Oso Dam, there are at least three different terrestrial to marine units exposed, these being, from oldest to youngest: the Vaqueros/Sespe, Topanga, and Monterey formations. Most of the marine mammals collected by the NHMLA crew come from the middle one, the Topanga Formation. This formation was deposited in shallow marine environments and has been dated to between 16.5-14.5 million years ago. The fauna of this formation is remarkable for its similarity to that of the Sharktooth Hill (STH) Bonebed in Bakersfield, CA, which is not entirely surprising, given the overlap in age. However, there are some notable differences between the faunas, including the absence of otariids in STH. In the Topanga Formation there are at least two early otariids, both belonging to the same genus, Eotaria crypta described a couple of years ago by Boessenecker & Churchill (2015), and Eotaria citrica, described in my new paper.
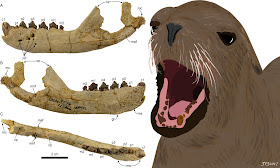 |
| FIGURE 2. On the left, the holotype and only known mandible of Eotaria citrica, in lateral, medial and occlusal views. On the right, a reconstruction of what this species may have looked like. |
(There are some useful explanations for some of these terms here.)
 |
| FIGURE 3. Evolutionary relationship between Eotaria spp. and otariids (from Velez-Juarbe, 2017:fig. 5). |
The second part of the paper, is an overview of the pinniped faunas of Southern California over the last 25 million years. This part was mainly the result of curiosity about the marine mammals of California, and also going over the collections over and over again. As I learned more both from the literature, looking at specimens and talking to colleagues, I thought this might be a story worth telling in more detail than had been done previously. The results, which are summarized in Figure 7 of the paper and the one below, are quite interesting. For starters, pinnipeds started out small, then got larger over time, which is something previous authors have noticed (e.g. Churchill et al., 2015) and that we see in other groups of marine organisms as well (Vermeij, 2012; Pyenson & Vermeij, 2016).
 |
| FIGURE 4. Overview of the pinniped faunas of Southern California over the last 25 million years. Each outline represents a distinct species. The present fauna is represented, left to right by: harbor seal (Phoca vitulina), northern fur seal (Callorhinus ursinus), California sea lion (Zalophus californianus), and northern elephant seal (Mirounga angustirostris) (modified from Velez-Juarbe, 2017:fig. 7). |
Beginning at around 16 million years ago, these faunas were composed by otariids, odobenids (walruses) and members of an extinct group called Allodesmus. Otariids represented the smallest bodied pinnipeds, while odobenids occupied mid-size ranges and Allodesmus were the largest bodied pinnipeds in those faunas. But that changed about 12-10 million years ago, when Allodesmus seems to have become extinct, at least in Southern California and walruses took over. It is then that odobenids diversified, with species occupying the mid to large body size ranges in the faunas. But things changed again, in relative recent times (less than 1.5 million years ago), when walruses became locally extinct and we see the appearance of true seals, like the harbor seals and northern elephant seal. These changes in the faunas, when a species is replaced by another, is what we refer to as faunal turnovers, and are usually tied, or can be correlated with local or global tectono-climatic events.
Nevertheless, there is still much more to be done with respect to the pinnipeds of Southern California, specially when it comes to their diversity as there seems to be more undescribed species in collections. The upcoming years will see more projects by colleagues (and myself too) where we'll continue to unravel the evolutionary history of pinnipeds and other marine tetrapods in the North Pacific region. So stay tuned!!!
References
Boersma, A. T., and N. D. Pyenson. 2016. Arktocara yakataga, a new fossil odontocete (Mammalia, Cetacea) from the Oligocene of Alaska and the antiquity of Platanistoidea. PeerJ 4:e2321; DOI 10.7717/peerj.2321
Boessenecker, R. W., and M. Churchill. 2015. The oldest known fur seal. Biology Letters 11(2):20140835; DOI 10.1098/rsbl.2014.0835
Churchill, M., M. T. Clementz, and N. Kohno. 2015. Cope's rule and the evolution of body size in Pinnipedimorpha (Mammalia: Carnivora). Evolution 69(1):201-215.
Pyenson, N. D., and G. J. Vermeij. 2016. The rise of ocean giants: maximum body size in Cenozoic marine mammals as an indicator for productivity in the Pacific and Atlantic Oceans. Biology Letters 12(7):20160186; DOI 10.1098./rsbl.2016.0186
Pyenson, N. D., J. Velez-Juarbe, C. S. Gutstein, H. Little, D. Vigil, and A. O'Dea. 2015. Isthminia panamensis, a new fossil inioid (Mammalia, Cetacea) from the Chagres Formation of Panama and the evolution of 'river dolphins' in the Americas. PeerJ 3:e1227; DOI 10.7717/peerj.1227
Velez-Juarbe, J. 2017. Eotaria citrica, sp. nov., a new stem otariid from the "Topanga" Formation of Southern California. PeerJ 5:e3022; DOI 10.7717/peerj.3022
Velez-Juarbe, J., A. R. Wood, C. De Gracia, and A. J. W. Hendy. 2015. Evolutionary patterns among living and fossil kogiid sperm whales: evidence from the Neogene of Central America. PLoS ONE 10(4):e0123909; DOI 10.1371/journal.pone.0123909
Vermeij, G. J. 2012. The evolution of gigantism on temperate seashores. Biological Journal of the Linnean Society 106(4):776-793; DOI 10.1111/j.1095-8312.2012.01897.x
No comments:
Post a Comment