Hace unos viernes atrás, salió mi
más reciente publicación. En la misma describo un nuevo género y especie de odontoceto (cetáceos dentados) proveniente de depósitos del Oligoceno tardío, entre 30.5-26.5 millones de años atrás, del estado de Washington. Este nuevo odontoceto al cual he bautizado como
Olympicetus avitus, se diferencia de sus parientes modernos por tener la nariz anterior a la órbita, dentición heterodonte (dientes diferenciados), una placa infraorbital de la maxilla, un cráneo alargado con amplia exposición del hueso parietal, maxilares que se extienden solo hasta la mitad anterior de la órbita y región temporal abierta dorsalmente, entre otras. La descripción de esta nueva especie es basada en dos especímenes, un neonato (LACM 126010) y un subadulto (LACM 149156) que fueron colectados de la misma localidad a lo largo de la Quebrada Murdock en el Condado de Clallam, Washington. Ambos especímenes estuvieron cubiertos por sedimentos muy duros de la Formación Pysht lo cual requirió que fuesen preparados con ácido, lo cual puede ser un reto, pero que si se hace correctamente resulta en especímenes casi perfectamente preparados y listos para ser estudiados.
 |
Comparación de algunas características que distinguen a Olympicetus de sus parientes actuales.
Arriba: los dos cráneos de Olympicetus avitus, el neonato (izquierda) y el subadulto y holotipo (derecha), desafortunadamente a ambos cráneos les falta parte del rostro.
Abajo: cráneo y mandíbulas de un Lagenorhynchus obliquidens, un pariente actual de Olympicetus. |
El
holotipo de
Olympicetus incluye una buena cantidad de dientes, los cuales demuestran que esta especie tenía dientes heterodontes, eso quiere decir que los dientes estaban diferenciados a lo largo de la dentadura. Esto es una característica heredada de otros cetáceos ancestrales, como contraste, la mayoría de odontocetos modernos tienen una dentadura homodonte, o sea, que todos los dientes son iguales y no puedes distinguir anteriores de posteriores. Desafortunadamente los dientes de
Olympicetus se habían caído durante el proceso de fosilización, y la posición exacta de cada diente solo se pudo estimar comparando con otros odontocetos con dentaduras heterodonte, como por ejemplo
Xenorophus sloani Kellogg, 1923, del Oligoceno temprano de Carolina del Sur.
 |
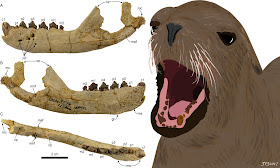
| Fragmento de mandíbula y los dientes del holotipo (LACM 149156) de Olympicetus avitus. Los dientes son heterodontes, y se pueden diferenciar en canino (A, P), premolariformes anteriores (D, E, Q, R), premolariformes posteriores (F-L, S-Y) y molariformes (M-O, Z-Bb) (modificado de la Figura 7 de Velez-Juarbe, 2017). |
El Oligoceno es una época importante para entender mejor la historia evolutiva de los odontocetos, ya que hacen su aparición en el registro fósil durante el principio de la misma hace unos 33.9 millones de años atrás. Interesantemente, al menos algunos grupos de esos odontocetos primordiales (i.e. los Xenorophidae) ya presentan muchas características que comparten con especies actuales, siendo la más relevante la
ecolocalización (Churchill et al., 2016; Park et al., 2016). La presencia de odontocetos en depósitos del Oligoceno en la región del Noroeste del Pacífico es bien conocida entre los paleontólogos ya que ha sido mencionado en la literatura científica durante los últimos 40 años (e.g. Whitmore & Sanders, 1977; Barnes et al., 2001). Sin embargo, solo dos especies habían sido descritas previo a
Olympicetus, ambas perteneciente a grupos distintos. De estos el más reciente que fue descrito es el allodelphinid
Arktocara yakataga, Boersma & Pyenson, 2016,
proveniente de depósitos del Oligoceno de Alaska. Los allodelphinids son un grupo de odontocetos de rostro alargado que incluye las especies:
Allodelphis pratti,
A. woodburnei,
Goedertius oregonensis,
Ninjadelphis ujiharai,
Zarhinocetus donnamatsonae y
Z. errabundus (Boersma & Pyenson, 2016; Kimura & Barnes, 2016). Este grupo en particular está actualmente extinto, pero están relacionados al
Delfín del Ganges (
Platanista gangetica) el cual es uno de los pocos delfines de río actuales y el último sobreviviente de un grupo que fue mucho más diverso en el pasado.
 |
| Vista dorsal de Goedertius oregonensis (LACM 123887) del Mioceno temprano de Oregon. Aquí podemos observar el rostro alargado que distingue a esta especie y otras relacionadas a la misma. |
El otro odontoceto del Oligoceno del Noroeste del Pacífico es
Simocetus rayi Fordyce, 2002. Esta especie, descrita en 2002, se considera suficientemente distinta para estar clasificada en su propia familia (Simocetidae).
Simocetus se distingue por su rostro relativamente corto y un poco agachado, dientes con bastante separación entre ellos, la nariz localizada anterior a la órbita y otras características que lo definen como un odontoceto basal. Además de compartir una región geográfica, algunas de las características que distinguen a
Simocetus se observan también en
Olympicetus y originalmente yo consideré que quizás pertenecían a la misma familia. Sin embargo, los resultados del análisis filogenético en mi estudio indican que
Olympicetus pertenece a un grupo distinto (ver filogenia abajo).
 |
| Relación filogenética de Olympicetus con otros odontocetos (modificado de la Figura 9 de Velez-Juarbe, 2017). |
Las relaciones entre estas especies podría cambiar si se descubre material más completo de
Olympicetus y/o cuando más especies parecidas a
Olympicetus y
Simocetus sean descritas y analizadas. Esto fue algo que sugerí en una presentación que di en la reunión anual de la Sociedad de Paleontología de Vertebrados en 2015, en ese entonces presenté unos resultados preliminares donde Simocetidae era una familia más diversa, quizás tanto como los Xenorophidae (e.g. Geisler et al., 2014; Boessenecker et al., 2017). También queda por ver si
Olympicetus y
Simocetus eran capaces de ecolocalización como los xenorophids. Existe material parecido en varias colecciones, así que el tiempo dirá!
References
Barnes, L. G., J. L. Goedert, and H. Furusawa. 2001. The earliest known echolocating toothed whales (Mammalia; Odontoceti): preliminary observations of fossils from Washington State. Mesa Southwestern Museum Bulletin 8:92-100.
Boersma, A. T., and N. D. Pyenson. 2016.
Arktocara yakataga, a new fossil odontocete (Mammalia, Cetacea) from the Oligocene of Alaska and the antiquity of Platanistoidea. PeerJ 4:e2321. doi: 10.7717/peerj.2321
Boessenecker, R. W., D. Fraser, M. Churchill, and J. H. Geisler. 2017. A toothless dwarf dolphin (Odontoceti: Xenorophidae) points to explosive feeding diversification of modern whales (Neoceti). Proceedings of the Royal Society B 284:20170531.
Churchill, M., M. Martinez-Caceres, C. de Muizon, J. Mneckowski, and J. H. Geisler. 2016. The origin of high-frequency hearing in whales. Current Biology 26:1-6.
Fordyce, R. E. 2002.
Simocetus rayi (Odontoceti: Simocetidae, New Family): a bizarre new archaic Oligocene dolphin from the Eastern Pacific. Smithsonian Contributions to Paleobiology 93:185-222.
Geisler, J. H., M. W. Colbert, and J. L. Carew. 2014. A new fossil species supports an early origin for toothed whale echolocation. Nature 508:383-386.
Kellogg, R. 1923. Description of an apparently new toothed cetacean from South Carolina. Smithsonian Contributions to Knowledge 76(7):1-7.
Kimura, T., and L. G. Barnes. 2016. New Miocene fossil Allodelphinidae (Cetacea, Odontoceti, Platanistoidea) from the North Pacific Ocean. Bulletin of the Gunma Museum of Natural History 20:1-58.
Park, T., E. M. G. Fitzgerald, and A. R. Evans. 2016. Ultrasonic hearing and echolocation in the earliest toothed whales. Biology Letters 12:20160060.
Velez-Juarbe, J. 2017. A new stem odontocete from the late Oligocene Pysht Formation in Washington State, U.S.A. Journal of Vertebrate Paleontology, DOI:10.1080/02724634.2017.1366916
Whitmore, F. C., Jr., and A. E. Sanders. 1977. Review of the Oligocene Cetacea. Systematic Zoology 25:304-320.